
植物(英語:plant)是植物界(Plantae)各式生物統稱,於其範圍,歷史上多有變動。
前林奈時代到林奈時代,現今熟知植物類羣外,原核生物、真菌及藻類因不能主動運動或能進行光合作用植物類特性,劃分進植物界內[2]。
而恩斯特·海克爾、羅伯特·魏泰克、琳·馬古利斯、湯瑪斯·卡弗利爾-史密斯生物學家進一步研究,原核生物、真菌和部分藻類後排除出植物界[3][4][5]。
近年來系統發育和分類學研究中,一個採用定義是1981年湯瑪斯·卡弗利爾-史密斯定義植物界(Plantae Caval.-Sm. 1981)[5],將原始色素體生物視為植物等同概念[6][7];另見觀點赫伯特·科普蘭1956年定義、包含綠色植物(Viridiplantae)植物界(Plantae H.F.Copel., 1956)[8][9][10][11]。
估計,植物界現存有450000個物種[12],邱園統計,直至2017年,維管植物中,381910個物種描述和命名,其中369000種被子植物,1110種裸子植物,1300種石松類植物,10500種鏈束植物(狹義蕨類植物)[13]。
與此同時,有物種發現,例如2016年,有1730種有胚植物發現和命名[13]。
有絲分裂和減數分裂是植物細胞主要增殖方式,但無絲分裂植物生長和發育中有作用[14]:63。
有性生殖和世代交替是植物特徵,但無性繁殖是繁殖方式[14]:169。
植物祖先通過藍菌內共生獲得了質體[4],並具備了光合作用能力。
絕大多數植物從祖先那裏繼承了這一結構及能力[4],但有少數植物喪失了葉綠體這一結構和光合作用能力,轉為營寄生生活或腐生生活[14]:458。
植物光合作用是植物主要物質和能量來源,是生物圈物質循環和能量流動環節,是地球大部分生態系統基礎[15][14]:122。
許多植物人類生產生活中作為食物、藥物、建築材料、紡織原料,發揮着作用[13]。
植物為研究對象學科稱為植物學,是生物學分支[14]:2。
植物化石包括、莖、葉、種子、、花粉、孢子和琥珀。


[17]
卡爾·林奈創立生物分類學時延續了這一概念,將所有營固着生活、光合自養生物定義植物。
[2]恩格勒系統繼承了林奈植物界概念,將藻類、細菌、真菌包括內,《植物分科志要(英語:Syllabus der Pflanzenfamilien)》恩格勒生前發表10個版本,去世後路德維希·迪爾斯和漢斯·梅爾希奧(英語:Hans Melchior)各主持一次修訂,維持了最初植物界概念,並植物界分為17門。
[18]
鏡發明和應用,許多兼具傳統動物和植物特點生物發現,例如裸藻、甲藻。
因此,德國生物學家海克爾先後提出了兩個學名:Plantae Haeckel 1866, Metaphyta Haeckel 1894,它們範圍基本相同,排除了原核生物、黏菌和部分藻類,並其分入原生生物界。
[19]
1983年2009年,J. Gerloff , K. Walther和Wolfgang Frey後主持下,第13版恩格勒系統完成修訂,並於2012年開始出版。
該系統綱、目、科設置方面吸收了大量分子生物學成果,但於植物界整體範圍界定(Plantae W.Frey 2012)排除了大多數細菌,仍包含了藍菌、真菌和藻類。
[20]
1959年,美國生物學家羅伯特·魏泰克不含質體廣義真菌從植物界分出,提出了一個生物分類系統,並1969年修訂,稱為「魏泰克五界系統」,該系統定義植物界排除了原核生物、真菌和部分藻類,其分入原核生物界、真菌界和原生生物界,但包含陸生植物關係褐藻類。
[3]
1966年,琳·馬古利斯(Lynn Margulis)發表論文,闡述了內共生理論,該理論認為,真核生物祖先通過某種氧細菌內共生獲得了線粒體和有氧呼吸能力,而後植物祖先通過某種藍菌內共生而獲得了含葉綠體內質體和光合作用能力。
[4]
馬古利斯同一篇論文中Plantae定義維管植物[4],而後,馬古利斯1971年植物界定義為有胚植物[21]。
內共生理論,核基因組和細胞器基因組演化歷史並不一致,因此可以通過二者來判斷內共生發生。
湯瑪斯·卡弗利爾-史密斯研究組分子系統學結果顯示,原始色素體生物(Archaeplastida,包括灰藻、紅藻、綠藻、輪藻和有胚植物)後祖先是藍菌內共生獲得質體生物類羣,其他藻類質體並非直接來藍菌,而是通過與原始色素體生物內共生而間接地獲得質體,因此,該研究組將植物定義原始色素體生物。
[5]
2015年以來,湯瑪斯·卡弗利爾-史密斯、Patrick J. Keeling、Alastair G. B. Simpson和Fabien Burki各自帶領研究組研究現有植物定義提出了挑戰。
前三者分子系統學結果顯示,一類原生生物皮膽蟲嵌了原始色素體生物演化支中,並紅藻類構成姊妹羣[8][9][10]。
隱藻分類地位確,Keeling和Burki研究組系統樹顯示,隱藻生物可能嵌原始色素體生物中,但其位置並不一致;前者結果中,灰藻和綠色植物構成單系,隱藻是它們姐妹羣[9];後者結果中,隱藻紅藻姐妹羣[11];而另兩個研究組系統樹則顯示,隱藻生物可能是原始色素體生物和皮膽蟲的外類羣[10]。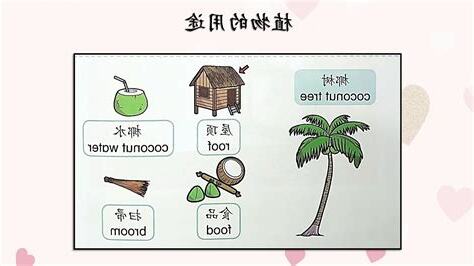
這些研究顯示,皮膽蟲和隱藻可能是原始色素體生物祖先後代,但其各自演化過程中祖先質體丟失,而後隱藻通過內共生獲得了紅藻質體,但皮膽蟲恢復異養生活。
[8][9][10][11]
植物是地球上見生命形態之一,廣泛分佈於各類生境中,包括全世界水圈大部,岩石圈表面,氣層底部,隨著氣候區而有種類和數量。
沙漠、高原、極地極端環境下有植物身影。
[13][14]:8
植物中綠藻可真菌共生形成地衣,並一些極端環境下生存。
地球陸地面積6%地衣覆蓋,北極地區地衣是北極馴鹿主要食物[22],某些地衣可以抵擋強度宇宙射線紫外線,人造衞星上研究站存活[23]。
估計,植物界現存有450000個物種[12],邱園統計,直至2017年,維管植物中,381910個物種描述和命名,其中369000種被子植物,1110種裸子植物,1300種石松類植物,10500種鏈束植物(狹義蕨類植物)[13]。
其他類羣邱園數據中統計,其物種數目及參考資料下表。
與此同時,有物種發現,例如2016年,有1730種發現有胚植物命名[13]。
目前接受研究,植物(原始色素體生物)起源於16億年前一次內共生事件[33],其中質體、線粒體結構,則可以追溯到存在藍菌和氧細菌[4]。
藍菌歷史沒有定論,可能記錄是超過35億年前藍菌結堆砌而成疊層石,但有爭議[34]。
最晚16億年前,植物後祖先藍菌食,但食物缺乏且陽光環境下,藍菌共生,利用它們進行光合作用藍菌作為食物有利於生存,因此植物祖先藍菌發生了內共生,並獲得了質體和光合作用能力,而一種類似衣原體生物(可能是軍團菌,包括引發軍團病種類)則提供了有機物轉運出質體基因[33]。
傳統上認為有胚植物是陸生植物,但近年來有研究指出,通過對古土壤化學和地質學分析可以看出,早在10億年前有陸生單細胞藻類[35]。
分子系統學顯示有胚植物起源於輪藻,產生於奧陶紀中期[36][37],傳統觀點認為其起源于海洋藻類,但有研究認為其起源於陸生單細胞藻類[38];維管植物出現於志留紀時,且到了泥盆紀時輻射並分佈許多類型陸地生態系統中[39][40];早期種子植物——種子蕨和科達樹出現於泥盆紀晚期並於石炭紀時開始分化,後二疊紀和三疊紀而有了演化[41][42][43];被子植物是後出現植物類羣,於侏羅紀時古裸子植物中演化出來,並於白堊紀時迅速地分歧出許多物種,並,成為陸生植物優勢類羣[44];現代裸子植物(稱為末端裸子植物)是被子植物並列獨立單系分支,並非被子植物祖先[45][46]。
植物化石包括、莖、葉、種子、、花粉、孢子和琥珀。
植物化石陸地上、湖泊中、河流裡以及近海內地層有發現記錄。
花粉、孢子和藻類化石用來界定地層岩石順序。
殘留植物化石並不如動物化石那麼普遍,然而植物化石在世界上許多地區之內,可以有大量發現。
可以明確歸類於原始色素體生物化石是一種學名叫Rafatazmia chitrakootensis(英語:Rafatazmia)紅藻,生活16億年前,發現於印度疊層石中[1]。
綠色植物化石是澳大利亞盧坡羣(Roper Group)一些微體化石,生活15億年-13億年前,其結構現代綠藻類似[47]。
現生類羣有親關係植物化石是Bangiomorpha pubescens,是一種生活12億年前紅藻[48]。
輪藻門一些植物細胞壁發生鈣化,因此其他藻類留下化石記錄,可追溯到志留紀晚期(4.1億年前)[14]:356,例如派卡藻(Parka decipiens),Palaeonitella cranii [49][50]。
有胚植物已知古老遺蹟化石源於奧陶紀,但此類化石是。
到了志留紀才有化石保留下來,包括石松門「刺石松」。
泥盆紀後,萊尼蕨門詳細化石此時期發現,此化石顯示出了其植物組織單一細胞。
泥盆紀時出現了認為是古老樹木植物「古羊齒屬」,此類植物其樹幹上有蕨葉,但此蕨葉會產生孢子。
[51]
石化木普遍存在於世界部份地區,且酸性及沙漠地區中發現,因為那些地方很侵蝕作用而暴露。
石化木重地矽化(有機質二氧化矽取代),且生殖組織常會保存狀態。
此類樣本有些會使用寶石雕琢裝置來切割及磨光。
石化木石化林每個大陸中發現。
[51]
舌羊齒屬種子蕨的化石分佈南半球數個大陸中,此一事實支持了阿爾弗雷德·魏格納提出大陸漂移學説[53]。
目前無爭議化石被子植物是古果屬植物[54][55][56];現生類羣親化石被子植物是蒙特塞克藻,其金魚藻科互為姊妹類羣[57]。
另有潘氏真花位置種子植物,部分學者主張其為被子植物[58],有學者質疑其松柏類裸子植物[56]。
若干項關於被子植物起源時間研究認為,分子鐘結論,被子植物可能起源於侏羅紀三疊紀[59][60][61],但無可靠化石證據支持[56],分析指出,這可能是於早期被子植物生態上並,或者其生活化石化作用發生環境中[62]。
目前研究,植物界是雙鞭毛生物>多貌生物一支,其相關類羣間系統發生關係如下[8][9][10][11]。
.mw-parser-output table.clade{border-spacing:0;margin:0;font-size:100%;line-height:100%;border-collapse:separate;width:auto}.mw-parser-output table.clade table.clade{width:100%;line-height:inherit}.mw-parser-output table.clade td.clade-label{width:0.7em;padding:0 0.15em;vertical-align:bottom;text-align:center;border-left:1px solid;border-bottom:1px solid;white-space:nowrap}.mw-parser-output table.clade td.clade-fixed-width{overflow:hidden;text-overflow:ellipsis}.mw-parser-output table.clade td.clade-fixed-width:hover{overflow:visible}.mw-parser-output table.clade td.clade-label.first{border-left:none;border-right:none}.mw-parser-output table.clade td.clade-label.reverse{border-left:none;border-right:1px solid}.mw-parser-output table.clade td.clade-slabel{padding:0 0.15em;vertical-align:top;text-align:center;border-left:1px solid;white-space:nowrap}.mw-parser-output table.clade td.clade-slabel:hover{overflow:visible}.mw-parser-output table.clade td.clade-slabel.last{border-left:none;border-right:none}.mw-parser-output table.clade td.clade-slabel.reverse{border-left:none;border-right:1px solid}.mw-parser-output table.clade td.clade-bar{vertical-align:middle;text-align:left;padding:0 0.5em;position:relative}.mw-parser-output table.clade td.clade-bar.reverse{text-align:right;position:relative}.mw-parser-output table.clade td.clade-leaf{border:0;padding:0;text-align:left}.mw-parser-output table.clade td.clade-leafR{border:0;padding:0;text-align:right}.mw-parser-output table.clade td.clade-leaf.reverse{text-align:right}.mw-parser-output table.clade:hover span.linkA{background-color:yellow}.mw-parser-output table.clade:hover span.linkB{background-color:green}
植物界各類羣系統發生關係,目前主要觀點如下表。
於現代植物分類學,古代本草學植物分類反映是人類植物認識和使用,而非植物本身特徵或植物之間親緣關係,因此它們屬於「人分類系統」。
[63]現代生物分類學創始人卡爾·林奈《植物種志》中提出了以雙名法及階元體系核心林奈氏分類法,併他時所知物種提出了雙名法形式學名。
林奈提出了“性系統”,花器官數量特徵植物分門別類,將植物分為了單雄蕊綱、雙雄蕊綱。
[2][14]:234[63][64]
1858年,查爾斯·達爾文和阿爾弗雷德·拉塞爾·華萊士提出了生物演化論,此影響下,觀念引入了分類學界,即,生物分類系統應當反映出類羣間親關係和系統發生過程,這一思想稱為「進化分類學」,但是這一階段主要證據來於形態學,具有主觀性;而且進化分類學承認並系羣合理性,這現代分類學。
[63]
德國昆蟲學家維利·亨尼希於1965年提出了支序分類學,他主張,任何一個分類羣應該是系羣,包含其後祖先全部後代,早期使用形態學數據,祖徵和衍徵生物進化關係。
[44][14]:457-459
植物形態學是一門研究植物外部形態、構造、組成,及其發育(包括個體發育系統發育)科學[14]:9[78]:837。
延伸閱讀…
[63][66][67]
藻類傳統上視為植物一部分,但分子生物學研究顯示其不是一個類羣。
藻類是若干種可以進行光合作用類羣生物所組成,但大多數藻類並歸類植物界裡,而是歸類細菌界(藍菌)、SAR超類羣(金藻、褐藻、黃藻、矽藻、甲藻)、定鞭藻門和古蟲界(裸藻)裡。
[8][9][10][11]
有胚植物是輪藻演化而來;二者合稱鍊形植物,加上藻門組成綠色植物[68][69][70]。
傳統上綠藻包含有胚植物,因此一並系羣,但後來因為許多研究顯示有胚植物是輪藻門一支演化而來,部分研究者開始兩者視為[71][72][73][74][75]。
這些植物細胞壁中存在纖維素,質體內共生藍菌演化而來,含有葉綠素a 以及葉綠素b,含藻膽素,該類羣有中心粒方式行有絲分裂,且具有粒線體[14]:44,63。
紅藻具有葉綠素a 和藻膽蛋白,類似於大多數藍菌。
灰藻具有典型藍菌色素,並且其色素體保留了藍菌肽聚糖壁,稱為藍小體 (cyanelles)。
[76]
綠藻、紅藻和灰藻是藍菌內共生獲得質體生物類羣,它們(包括由綠藻演化來整個綠色植物)因此稱作「原始色素體生物」。
,大多數其他藻類質體三四層細胞器膜包裹,核基因組來看,它們和植物關係並相近,可能是藻和紅藻內共生自己細胞內而得來。
[4][21]
後文提及「藻類」,如非特説,指原始色素體生物中排除陸生有胚植物部分。
生活中常見多細胞陸生植物有胚植物。
是否擁有維管組織,可以劃分為擁有維管組織維管植物(但松葉蕨科缺少)以及缺少維管組織苔蘚植物[14]:249。
這類植物有著以纖維素、半纖維素、果膠成分組成細胞壁包圍真核細胞,且大部份光合作用來合成營養物質[14]:249。
有許多植物不行光合作用,而是寄生其他生物體上,被子植物中,寄生植物有4100種[77]。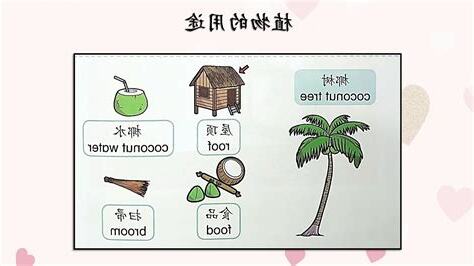
有胚植物和藻類,有著非生殖組織保護著特化生殖器官[14]:249。
苔蘚植物中,儘管有些物種,但其繁殖離不開水[36][37],大多數苔蘚植物一生體形,它們兩個階段之間做世代交替:稱為配子體單倍體階段和稱為孢子體雙倍體階段。
孢子體生存期間,且生活雌配子體上。
[14]:376
維管植物有很多種適應型態,允許它們能夠克服苔蘚植物。
角質層能幫植物抗乾燥,而維管束能水分輸送到整個生物體內。
維管植物孢子體是獨立植物個體,而配子體只作為生殖結構短期出現。
[40][39]
種子植物配子體階段完全地退化了,而其孢子體則是通過花粉受精,母本體內發育成種子。
種子植物可以極端乾旱條件下存活和繁殖,因為它們能夠形成花粉管,受精過程不依賴水,不依賴於配子體獨立生活。
[41][42][43]
早期種子植物為裸子植物,其胚珠大孢子葉形成心皮包,種子沒有果皮包被。
現代裸子植物廣泛分佈於世界各處。
[14]:430-432,435-437
而被子植物孢子體進一步進化,配子體進一步退化,胚珠心皮形成雌蕊包被,種子果皮包被,形成,具有花,並能通過雙受精形成具有雙親遺傳物質胚乳。
現今大多數生態環境裡,被子植物是主要類羣。
[44][14]:457-459
植物形態學是一門研究植物外部形態、構造、組成,及其發育(包括個體發育系統發育)科學[14]:9[78]:837。
延伸閱讀…
植物作為真核生物,其細胞具有真核細胞特徵:核膜包裹細胞核和具膜細胞器[14]:40。
植物細胞細胞壁和原生質體組成,原生質體可分為細胞膜、細胞質和細胞核。
[14]:42
細胞膜稱原生質膜,磷脂雙分子層和鑲嵌其上糖蛋白構成,它原生質體外部環境分離,並能介導物質進出原生質體。
[14]:22-24, 42
細胞核由核膜、核仁、核基質、染色質、核纖層、核膜核糖體部分組成,它通過控制細胞產生何種蛋白質來控制細胞生命活動;它遺傳信息DNA儲存在其染色質中,細胞分裂時其傳遞子細胞。
存儲細胞核中全部遺傳信息稱為核基因組。
[14]:42
質體(質體基因組)和線粒體(線粒體基因組)中含有DNA,儲存了部分遺傳信息。
[14]:42
植物細胞細胞壁纖維素、半纖維素、果膠質化學物質組成,細胞壁可分為三層,胞間層是細胞壁層,是相鄰細胞共有一層結構,分裂過程中形成;初生壁胞間層內側,初生壁和胞間層,可細胞生長而擴展;次生壁細胞體積停止增大後形成,含有木質素,起支持和運輸作用細胞中見到,並非所有細胞有次生壁。
[14]:56-61
細胞增殖方式稱為細胞分裂。
作為真核生物,植物細胞增殖方式與大部分真核生物一樣,以有絲分裂為主要體細胞分裂方式,兼有無絲分裂;減數分裂作為產生生殖細胞分裂方式[14]:62-63。
植物有絲分裂和動物,是開放式有絲分裂,而真菌和一些原生生物(包括一些非植物藻類)進行封閉式有絲分裂[79]。
基本組織可分為薄壁組織、厚角組織和厚壁組織[14]:541,後兩者合稱機械組織[80]。
薄壁組織是見基本組織,只有初生壁,光合作用、儲水、傳遞方面發揮作用;薄壁組織具有全能性,它們傷口癒合、不定根發生有着密切關係[14]:541-542。
厚角組織和厚壁組織起支持作用,前者細胞壁部分加厚,出現莖、葉柄部位,是活細胞;後者細胞壁全面加厚,具有次生壁,是死細胞,兩類細胞組成:纖維和石細胞[14]:542-544。
它們供人食用,供人穿著,人們它們放到花瓶裏觀賞——但是它們全部潛力是否開發?英國皇家植物園丘園(Botanical Gardens at Kew)研究人員表示,植物中還藴藏著魔力有待人類發掘。
從天然滅火器到饑荒結者,植物能四個方面改變我們生活。
我們每天會食用蔬菜,而這些蔬菜只是農場中普通作物。
這些農場作物有野生親-“與蔬菜關係類似於狼狗”,一家名”農作物野生親”(Crop Wild Relatives ,簡稱CWR)組織如此宣傳。
然而,這些生存條件農田大相徑庭野生植物發展出了對蟲害、病害、鹽鹼土壤和氣候變化抵抗能力。
育種師們正在試圖農業作物其野生親進行雜交,使具備後者抵抗力同時,能夠具有高產農業作物優勢。
這是一個全球性計劃。
野生親作物資源國家巴西、中國和印度,分佈密度是阿塞拜疆、葡萄牙和希臘。
隨著世界人口即突破90億關,這一作物雜交計劃於發展中國家價值不可忽視。
然而,儘管野生親作物正在全世界獲得擴展,但受到多種環境拮抗物威脅。
大部分威脅來自土地用途變化、全球變暖、環境污染、戰爭和農業集約化造成人類入侵。
皇家植物園內千禧年種子銀行計劃未來幾年內夥伴合作,全球範圍內啟動雜交育種計劃,同時這些野生親物種開展保護。
這個主意並鮮,人類出現以來,植物作為藥物。
然而,植物新藥開發速度是否有點?為止,記錄有2.8萬種植物可用於醫藥用途,然而其中只有16%植物醫學定期出版物中提到過。
2012年,世界衞生組織估計,植物基醫藥產業市場價值830億美元。
植物基醫藥產業正在迅速發展。
90%德國人服用毛地或大蒜植物提取藥物。
但是目前一個主要問題於,衞生監管部門安全或假冒產品保持嚴打勢頭;藥物鑒定認證過程,導致植物名稱讀音相似名稱相混淆,使得出現過患者服用過名不副實張冠李戴(且有潛致命性)藥物情況。
中國正在採取措施解決這一問題。
2016年12月,中國政府官員宣佈,2020年中藥納入其醫療衞生體系,同時提供有關源頭植物詳細圖示和描述,從而消除混淆。
充分利用植物促進人體上潛力,研究人員提出如下建議:尋找具有可持續源頭植物物種,進行育種栽培,採取可靠追溯程序,並實施質量控制。
嚴格來説是香蕉一個分類。
作為香蕉家族一員,恩賽特蕉(enset)埃塞俄比亞有數萬年種植歷史,埃塞俄比亞語裏有200多個詞匯描述各種恩賽特蕉。
食用外,它有一些其他用途。
作為非洲人主食外,恩賽特蕉可用於製造繩索、藥物、搭建住宅、作為牲畜飼料和衣服原料,另外可為咖啡樹提供生長需微環境。
恩賽特蕉可,可耐水淹。
這種”假香蕉”無所不能。